



 Éd. 1971-1976
Éd. 1971-1976Archæopteryx
Oiseau fossile le plus ancien connu. Il provient du Jurassique supérieur de Bavière (calcaire lithographique, étage kimméridgien).
Quatre exemplaires seulement de ce fossile ont été décrits : le premier, découvert en 1861 à Langenaltheimer Haardt, près de Pappenheim, fut acheté par sir R. Owen, pour le British Museum, 700 livres, somme énorme pour l’époque ; le deuxième, trouvé en 1877 près d’Eichstätt, fut vendu au musée de Berlin ; le troisième spécimen provient de la même localité que le premier et d’un horizon voisin ; interprété comme étant un Archœopteryx en 1958, il est en la possession de l’université d’Erlangen ; un quatrième, trouvé en 1867, mais longtemps mal interprété, a été identifié en 1970. L’étude précise de ces spécimens a montré qu’ils appartenaient tous à un seul genre Archæopteryx, mais à deux espèces différentes (A. lithographica du British Museum, A. Siemensi du musée de Berlin).
Caractères reptiliens
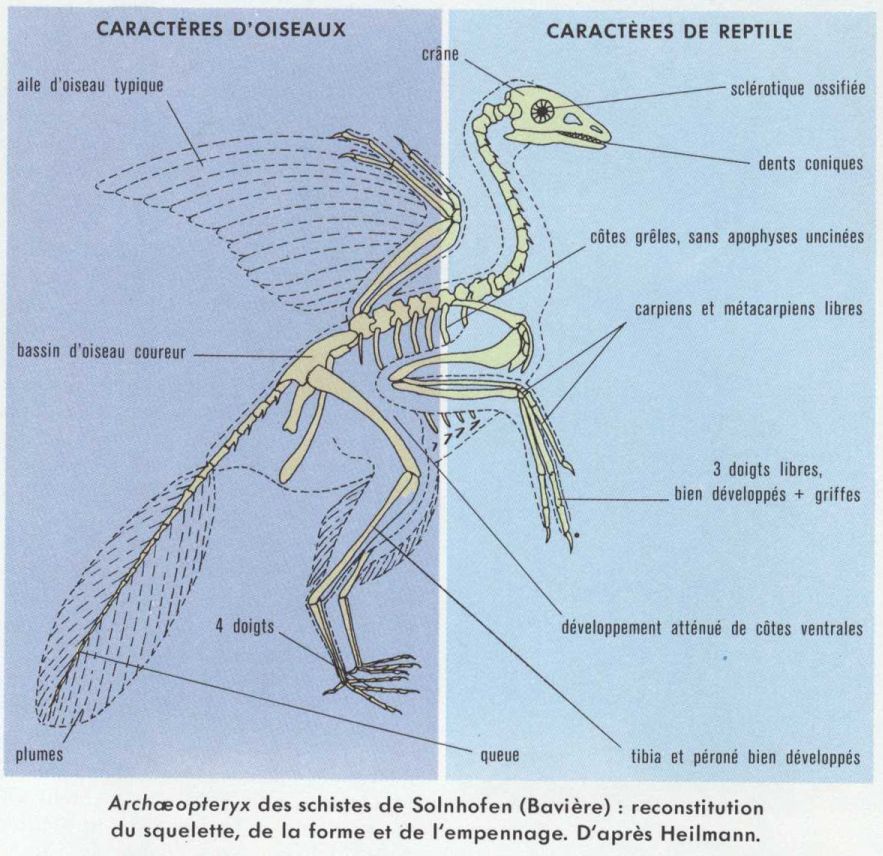
Archæopteryx est incontestablement un Oiseau, car il présente des plumes disposées comme chez les Oiseaux actuels ; de plus, les deux clavicules sont soudées en une fourchette. Mais Archæopteryx possède aussi de nombreux caractères archaïques et reptiliens ; ainsi, il présente une longue queue à vertèbres distinctes ; celles-ci sont biconcaves ; le sacrum ne comprend pas plus de six vertèbres, alors que le sacrum des Oiseaux est formé d’au moins onze vertèbres ; les mâchoires sont garnies de dents ; les métacarpiens sont totalement séparés (ils sont réunis entre eux à leur extrémité distale chez les Oiseaux et avec deux carpiens à leur extrémité proximale) ; les doigts sont terminés par des griffes ; les os métatarsiens sont toujours, au moins en partie, séparés les uns des autres sans formation d’un tarsométatarsien unique comme chez les Oiseaux (« os canon ») ; les côtes grêles, sans apophyses uncinées (apophyses insérées postérieurement sur chaque côte en direction de la côte postérieure), ne sont pas articulées avec le sternum ; il existe des côtes ventrales renforçant la paroi inférieure du corps.
Archæopteryx apparaît donc comme une forme intermédiaire entre Reptiles et Oiseaux, et le mélange de caractères observables sur ce fossile est une excellente preuve de l’évolution animale ; mais, si l’on considère seuls les caractères du squelette, on constate que celui-ci est nettement reptilien. Le même retard concerne aussi l’évolution de l’encéphale ; certes, l’encéphale d’Archæopteryx n’est pas connu, mais, sur l’exemplaire du British Museum, le moule interne du crâne est visible ; or, sur cette empreinte, la trace des lobes optiques est bien visible entre celle des hémisphères et celle du cervelet, tandis que, chez les Oiseaux, les lobes optiques sont en majeure partie recouverts. Les os longs des Oiseaux sont pneumatiques, c’est-à-dire qu’ils sont remplis d’air et communiquent avec des sacs, les sacs aériens de la cavité du corps, par des orifices (dispositif d’allégement de l’animal). On n’a jamais observé sur les os d’Archæopteryx de tels orifices ; les os de celui-ci n’étaient donc pas pneumatiques, mais ils possédaient néanmoins de grandes cavités médullaires. Les plumes présentaient un axe, le rachis, portant des barbes et des barbules. On distingue parmi elles des rémiges primaires, des rémiges secondaires (plumes des ailes insérées respectivement sur les os de la main et sur le cubitus) et des rectrices (plumes de la queue).
Archæopteryx et l’origine du vol
L’anatomie d’Archæopteryx montre que cet animal devait voler assez mal (absence de bréchet, cou peu mobile, musculature thoracique probablement assez peu active, lobes optiques et cervelet peu développés, etc.). Archæopteryx planait d’une branche à l’autre et, par ses griffes, devait pouvoir s’accrocher aux arbres ; en raison notamment de sa queue emplumée, il devait pouvoir difficilement s’écarter de sa trajectoire, à l’inverse des Oiseaux modernes. Comme, d’autre part, il avait des caractères d’arboricoles (présence de griffes, un doigt de pied opposable aux autres doigts) et non d’animal coureur (les plumes de la queue sont incompatibles avec un tel mode de vie), on peut en conclure que son vol est apparu non pas comme une adaptation à la course, mais comme une conséquence de la vie dans les arbres. Ainsi, en dehors de son intérêt anatomique (Archæopteryx a à la fois des caractères de Reptiles et d’Oiseaux), apporte-t-il aussi la solution du problème controversé de l’origine du vol.
J. P. L.
G. de Beer, Archæopteryx Lithographica ; a Study Based upon the British Museum Specimen (Londres, 1954).
archégone
Chez les Bryophytes et les plantes vasculaires, organe qui contient l’oosphère, ou gamète femelle.
Suivant les groupes, l’archégone est diversement développé, mais il présente partout les mêmes caractéristiques générales : il affecte la forme d’une amphore munie d’un col plus ou moins long, et c’est dans la partie renflée, ou ventre, que se trouve l’oosphère. De tels organes n’existent que chez les Bryophytes, les Ptéridophytes, les Gymnospermes, et certains auteurs lui trouvent des correspondants très simplifiés chez les Angiospermes. Chez les Algues et les Champignons, les enveloppes des oosphères ne sont pas cellulaires, mais membraneuses, et l’on parle alors d’oogones et non d’archégones. Cependant, les oogones des Characées apparaissent comme une amorce de ce que sont les archégones des plantes supérieures.
Bryophytes
Chez les Bryophytes, les archégones sont bien développés ; on les trouve à l’extrémité de certains rameaux feuilles au milieu d’un involucre. Ces organes se forment à partir d’une cellule superficielle qui se divise transversalement ; la cellule supérieure se cloisonne de nouveau pour donner quelques cellules pariétales et une cellule interne. Les cellules pariétales, en se multipliant et en se développant, deviendront d’une part la paroi du ventre de l’archégone (2 ou 3 couches de cellules) et d’autre part le col à paroi mince (une seule), mais parfois très long, jusqu’à 50 étages de cellules superposées. La cellule interne ou centrale se divise d’abord une fois, pour donner une cellule fille supérieure, qui est à l’origine des cellules du canal du col, puis une seconde fois ; de là proviendront la dernière cellule du canal vers le ventre et l’oosphère, ou gamète. Les cellules du canal se gélifient au moment où l’oosphère est fécondable et constituent un mucilage qui a le double rôle d’attirer les anthérozoïdes, surtout par diffusion de saccharose (chimiotropisme), et de permettre aux anthérozoïdes d’atteindre ensuite l’oosphère. Après la fécondation, l’oosphère s’entoure d’une membrane cellulosique, la paroi du ventre de l’archégone s’épaissit, le col se flétrit et l’œuf fécondé se développe à l’intérieur du ventre, nourri par les tissus de l’extrémité de la tige.
Plus tard, lorsque le sporogone (embryon à 2 n chromosomes) est formé, l’archégone se déchire, et l’on retrouve au sommet de la capsule du sporogone sa partie supérieure appelée coiffe.
Chez les Marchantias (Hépatiques), on observe des archégones appendus sous le chapeau fertile femelle. Leur structure est identique à celle des archégones des Mousses.