




cerveau (suite)
Embryologie et structure
Développement embryonnaire
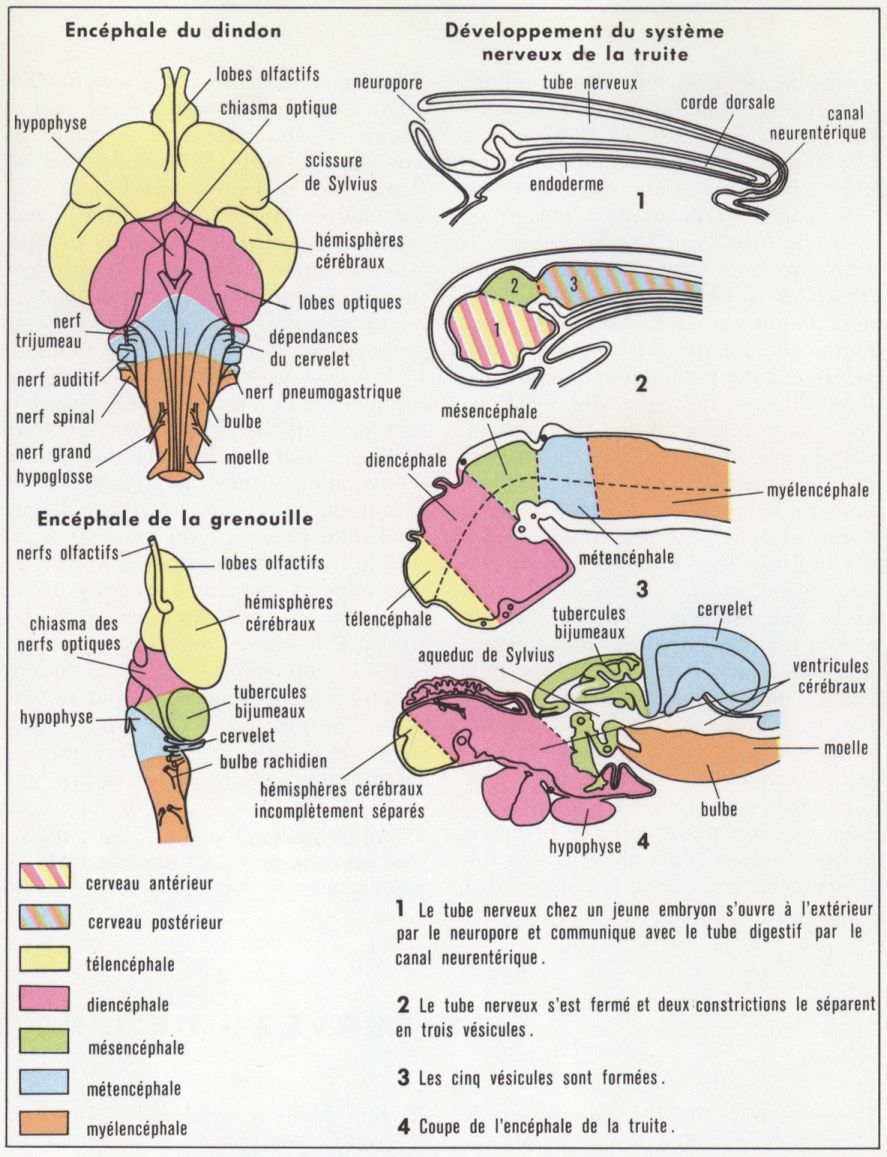
Le système nerveux est une formation d’origine ectodermique. Alors qu’il reste intra- ou basi-épithélial chez les animaux primitifs, chez les autres groupes il s’isole de l’ectoderme et forme des structures pleines (ganglions et cordons : Annélides, Mollusques et Arthropodes) ou un tube creux (Vertébrés). Dans ce dernier cas, la mise en place du système nerveux a lieu très tôt au cours du développement embryonnaire, juste après la mise en place des feuillets germinatifs ; c’est la neurulation. La portion dorso-médiane de l’ectoderme s’épaissit en une plaque neurale, qui se creuse ensuite en une gouttière nerveuse. Les bords de cette gouttière viennent se souder dorsalement et ferment ainsi le tube nerveux ; il subsiste pendant un temps très court un orifice antérieur (neuropore) et un orifice postérieur communiquant parfois avec l’intestin (canal neurentérique). La portion antérieure du tube nerveux se renfle en une vésicule encéphalique : la portion postérieure reste mince et rectiligne, et deviendra la moelle épinière.
La vésicule antérieure reste indivise chez les Procordés, notamment chez Amphioxus. Chez les Vertébrés, elle se subdivise, par des étranglements d’aspect métamérique, en trois vésicules, appelées, d’avant en arrière, prosencéphale, mésencéphale et rhombencéphale. Rapidement, la première et la dernière de ces vésicules se subdivisent à leur tour dans leur seule portion dorsale, si bien qu’il existe au total cinq vésicules encéphaliques, qu’on retrouve, bien que difficilement, chez l’adulte ; ces vésicules sont, d’avant en arrière, le télencéphale (hémisphères cérébraux et corps striés), le diencéphale (couches optiques et hypothalamus), le mésencéphale (toit optique et tegmentum), le métencéphale (cervelet) et le myélencéphale (tronc cérébral pro parte, moelle allongée ou bulbe rachidien chez les Mammifères).
Courbures, cavités, enveloppes
Au cours de la différenciation des diverses vésicules encéphaliques se produisent des flexures qui transforment le tube encéphalique rectiligne de l’embryon en un organe dont l’axe longitudinal est très altéré, notamment chez les Oiseaux et les Mammifères. Chez ces derniers, les flexures encéphalique et nucale courbent l’encéphale ventralement, tandis que les flexures pontique et cérébrale ont l’orientation inverse.
Ce sont les parois du tube nerveux qui fournissent la substance nerveuse ; la lumière est emplie du liquide céphalo-rachidien. Elle forme dans la moelle épinière un canal étroit, l’épendyme. Dans l’encéphale, elle constitue deux renflements, que sépare un pédoncule, l’aqueduc de Sylvius, ou canal mésencéphalique. Le renflement postérieur, ou 4e ventricule, est ventral au cervelet ; le renflement antérieur, ou 3e ventricule, est diencéphalique, et il communique par deux orifices latéraux, les trous de Monro, avec les ventricules latéraux, ou ventricules 1 et 2, situés dans chacun des hémisphères cérébraux. Des organes épithélio-vasculaires, les plexus choroïdes, situés au plafond des ventricules 3 et 4, sécrètent le liquide céphalo-rachidien.
L’encéphale est isolé du crâne par une enveloppe intermédiaire, formant les méninges. La portion externe, ectoméninge ou dure-mère, est d’origine mésenchymateuse ; entre les diverses vésicules, elle forme des plis rentrants : la tente du cervelet (entre cerveau et cervelet) et la faux du cerveau (entre les deux hémisphères), qui, de plus, peuvent s’ossifier, par exemple chez les Mammifères carnivores. La portion interne, ou leptoméninge, est d’origine nerveuse ; elle participe à la formation des plexus choroïdes. Chez les Mammifères, la leptoméninge se subdivise en pie-mère, au contact direct du tissu nerveux, et en arachnoïde, tissu mésenchymateux lâche et vascularisé.
« Substance blanche » et « substance grise »
Chez le jeune embryon, toutes les cellules nerveuses sont placées en bordure des ventricules. Elles subissent alors à la fois des mitoses (multiplications cellulaires) et des migrations qui ne laissent au contact du ventricule que les cellules épendymaires. Les autres cellules, concentrées dans la portion profonde du tube nerveux, y forment le manteau, ou pallium. Les cellules du pallium sont ou bien des cellules névrogliques. aux fonctions métaboliques diverses, ou bien des neurones, spécialisés dans la conduction de l’influx nerveux. Ces derniers se différencient et acquièrent des prolongements axoniques situés à la périphérie du pallium, si bien qu’à ce stade le tube nerveux contient une substance grise centrale (riche en neurones et en cellules gliales) qu’entoure la substance blanche périphérique (riche en fibres myélinisées). C’est la structure de la moelle épinière et du tronc cérébral. Aux niveaux des hémisphères cérébraux et du cervelet, la migration cellulaire est plus intense et porte les cellules au voisinage immédiat de la paroi externe du tube, y formant un cortex ; le cortex cérébelleux est présent chez tous les Vertébrés, tandis que le cortex cérébral n’apparaît que chez les plus évolués des Vertébrés, notamment chez les Mammifères.

Composants fonctionnels et nerfs crâniens
Disposition générale des voies nerveuses
Les neurones se groupent en cordons dans la moelle épinière et le tronc cérébral postérieur, en noyaux dans les régions sous-corticales et en nappes dans le cortex. Les fibres nerveuses s’assemblent elles-mêmes en faisceaux, ou tractus, en commissures et en décussations. Mais ces regroupements de fibres et de cellules ne se font pas « au hasard ». Les portions dorsales sont réservées aux centres sensoriels et de corrélation ; les portions ventrales, aux centres moteurs et de coordination. Dans la moelle épinière et le tronc cérébral, ces zones sont limitées par un sillon ventriculaire, le sillon limitant, qui sépare ainsi la lame alaire, dorsale et sensorielle, de la lame basale, ventrale et motrice. Il en va de même des fibres, qui sont afférentes ou sensitives dorsalement, efférentes ou motrices ventralement. De plus, au sein même de ces zones, existent des subdivisions elles aussi de nature fonctionnelle ; les centres sensoriels sont étages en trois niveaux : le niveau le plus dorsal reçoit les fibres d’origine cutanée ou extéroceptives, le niveau intermédiaire les fibres d’origine musculaire ou proprioceptives, et le niveau le plus profond les fibres sensorielles viscérales ou intéroceptives. De même, les centres moteurs comportent une portion profonde viscéro-motrice (dont les fibres empruntent le système nerveux autonome ou sympathique) et une portion externe somato-motrice, innervant la musculature striée d’origine somitique.